






Hücresel Solunum Nedir?

Fotosentez sürecinde, bitkiler ve diğer fotosentetik üreticiler, enerjiyi kimyasal bağlarında depolayan glikoz oluştururlar. Daha sonra, hem bitkiler hem de hayvanlar gibi tüketiciler, topluca hücresel solunum olarak adlandırılan bir dizi metabolik yoldan geçer. Hücresel solunum, glikozdaki bağlardan enerjiyi alır ve tüm canlıların kullanabileceği bir forma dönüştürür.
Öğrenme Hedefleri
- Glikoliz sürecini tanımlayın ve reaktanlarını ve ürünlerini tanımlayın.
- Piruvat oksidasyon sürecini tanımlayın ve reaktanları ve ürünlerini tanımlayın.
- Sitrik asit döngüsünün ( Krebs döngüsü ) sürecini tanımlayın ve reaktanları ve ürünlerini tanımlayın.
- Solunum zincirini ( elektron taşıma zinciri ) ve hücresel solunumdaki rolünü tanımlayın.
Hücresel solunum, tüm canlıların glikozu enerjiye dönüştürmek için kullandığı bir süreçtir. Ototroflar ( bitkiler gibi ) fotosentez sırasında glikoz üretir. Heterotroflar ( insanlar gibi ) glikoz elde etmek için diğer canlıları yutarlar. Süreç karmaşık görünse de, bu sayfa sizi hücresel solunumun her bir bölümünün temel unsurlarına götürür.
Glikoliz
Glikoliz, hücresel metabolizma için enerji elde etmek üzere glikozun parçalanmasındaki ilk adımdır. Neredeyse tüm canlı organizmalar, metabolizmalarının bir parçası olarak glikoliz gerçekleştirir. İşlem oksijen kullanmaz ve bu nedenle anaerobiktir ( oksijen kullanan işlemlere aerobik denir ). Glikoliz, hem prokaryotik hem de ökaryotik hücrelerin sitoplazmasında gerçekleşir. Glikoz heterotrofik hücrelere iki şekilde girer.
- Taşımanın glikoz konsantrasyonu gradyanına karşı gerçekleştiği ikincil aktif taşıma yoluyla.
- Glikoz taşıyıcı proteinler olarak da bilinen GLUT proteinleri adı verilen bir grup integral protein aracılığıyla. Bu taşıyıcılar, glikozun kolaylaştırılmış difüzyonuna yardımcı olur.
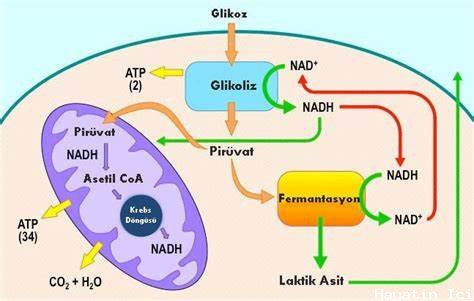
Glikoliz, tek bir glikoz molekülünün altı karbonlu halka şeklindeki yapısıyla başlar ve piruvat adı verilen üç karbonlu bir şekerin iki molekülü ile sona erer ( Şekil 1 ).

Glikoliz, iki ayrı yarıya bölünmüş on adımdan oluşur. Glikolizin ilk yarısı, enerji gerektiren adımlar olarak da bilinir. Bu yol, glikoz molekülünü hücrede yakalar ve onu değiştirmek için enerji kullanır, böylece altı karbonlu şeker molekülü iki üç karbonlu moleküle eşit olarak bölünebilir. Glikolizin ikinci yarısı ( enerji salma adımları olarak da bilinir ) moleküllerden enerji çıkarır ve onu NAD'nin indirgenmiş formu olan ATP ve NADH formunda depolar.
Glikolizin İlk Yarısı ( Enerji Gerektiren Adımlar )

Aşama 1. Glikolizdeki ilk adım, altı karbonlu şekerlerin fosforilasyonunu katalize eden geniş özgüllüğe sahip bir enzim olan hekzokinaz tarafından katalize edilir. Hekzokinaz, fosfat kaynağı olarak ATP kullanarak glikozu fosforile eder ve daha reaktif bir glikoz formu olan glikoz-6-fosfat üretir. Bu reaksiyon, fosforile glikoz molekülünün GLUT proteinleri ile etkileşime devam etmesini önler ve negatif yüklü fosfat, plazma zarının hidrofobik iç kısmını geçmesine izin vermeyeceği için artık hücreyi terk edemez.
Adım 2. Glikolizin ikinci adımında, bir izomeraz, glikoz-6-fosfatı izomerlerinden biri olan fruktoz-6-fosfata dönüştürür. Bir izomeraz, bir molekülün izomerlerinden birine dönüşümünü katalize eden bir enzimdir. Fosfoglukozdan fosfofruktoza olan bu değişim, şekerin nihai olarak iki üç karbonlu moleküle bölünmesine izin verir.
Aşama 3. Üçüncü adım, fosfofruktokinaz enzimi tarafından katalize edilen fruktoz-6-fosfatın fosforilasyonudur. İkinci bir ATP molekülü, fruktoz-6-fosfata yüksek enerjili bir fosfat bağışlayarak fruktoz-1,6-bifosfat üretir. Bu yolda, fosfofruktokinaz, hız sınırlayıcı bir enzimdir. ADP konsantrasyonu yüksek olduğunda aktiftir; ADP seviyeleri düşük ve ATP konsantrasyonu yüksek olduğunda daha az aktiftir. Bu nedenle, sistemde "yeterli" ATP varsa, yol yavaşlar. ATP, glikoz katabolizmasının son ürünü olduğu için bu bir tür son ürün inhibisyonudur.
Adım 4. Yeni eklenen yüksek enerjili fosfatlar, fruktoz-1,6-bifosfatı daha da dengesizleştirir. Glikolizdeki dördüncü adım, 1,6-bifosfatı iki üç karbonlu izomere bölmek için bir enzim olan aldolaz kullanır: dihidroksiaseton-fosfat ve gliseraldehit-3-fosfat.
Adım 5. Beşinci adımda, bir izomeraz, dihidroksiaseton-fosfatı izomeri olan gliseraldehit-3-fosfata dönüştürür. Böylece yol, tek bir izomerin iki molekülü ile devam edecektir. Yolun bu noktasında, bir glikoz molekülünün parçalanmasında iki ATP molekülünden net bir enerji yatırımı vardır.
Glikolizin İkinci Yarısı ( Enerji Açığa Çıkaran Adımlar )
Şimdiye kadar, glikoliz hücreye iki ATP molekülüne mal oldu ve iki küçük, üç karbonlu şeker molekülü üretti. Bu moleküllerin her ikisi de yolun ikinci yarısı boyunca ilerleyecek ve ilk yatırım olarak kullanılan iki ATP molekülünü geri ödemek için yeterli enerji çıkarılacak ve iki ek ATP molekülü ve iki daha yüksek enerjili NADH molekülü hücresi için bir kar üretecektir.

Adım 6. Glikolizdeki altıncı adım ( Şekil 3 ) şekeri ( gliseraldehit-3-fosfat ) oksitleyerek, elektron taşıyıcı NAD+ tarafından alınan ve NADH üreten yüksek enerjili elektronları çıkarır. Şeker daha sonra ikinci bir fosfat grubunun eklenmesiyle fosforile edilir ve 1,3-bifosfogliserat üretilir. İkinci fosfat grubunun başka bir ATP molekülü gerektirmediğine dikkat edin.
İşte yine bu yol için potansiyel bir sınırlayıcı faktör. Reaksiyonun devamı, elektron taşıyıcısının oksitlenmiş formu olan NAD+'nın mevcudiyetine bağlıdır. Bu nedenle, bu adımın devam etmesi için NADH'nin sürekli olarak NAD+'ya oksitlenmesi gerekir. NAD+ mevcut değilse, glikolizin ikinci yarısı yavaşlar veya durur. Sistemde oksijen mevcutsa, NADH dolaylı olarak da olsa kolayca oksitlenecek ve bu süreçte salınan hidrojenden gelen yüksek enerjili elektronlar ATP üretmek için kullanılacaktır. Oksijensiz bir ortamda, alternatif bir yol ( fermantasyon ) NADH'nin NAD+ 'ya oksidasyonunu sağlayabilir.
Adım 7. Yedinci adımda, fosfogliserat kinaz ( ters reaksiyon için adlandırılan bir enzim ) tarafından katalize edilen 1,3-bifosfogliserat, ADP'ye yüksek enerjili bir fosfat bağışlayarak bir ATP molekülü oluşturur. ( Bu, substrat düzeyinde fosforilasyonun bir örneğidir.) 1,3-bifosfogliserat üzerindeki bir karbonil grubu, bir karboksil grubuna oksitlenir ve 3-fosfogliserat oluşur.
Adım 8. Sekizinci adımda, 3-fosfogliseratta kalan fosfat grubu, üçüncü karbondan ikinci karbona hareket ederek 2-fosfogliserat ( 3-fosfogliseratın bir izomeri ) üretir. Bu adımı katalize eden enzim bir mutazdır ( bir tür izomeraz ).
Adım 9. Enolase dokuzuncu adımı katalize eder. Bu enzim 2-fosfogliseriatın yapısından su kaybetmesine neden olur; bu bir dehidrasyon reaksiyonudur ve kalan fosfat bağındaki potansiyel enerjiyi artıran ve fosfoenolpiruvat ( PEP ) üreten bir çift bağ oluşumuyla sonuçlanır.
Adım 10. Glikolizdeki son adım, piruvat kinaz enzimi tarafından katalize edilir ( bu durumda enzim, piruvatın PEP'e dönüşümünün ters reaksiyonu için adlandırılır ) ve substrat düzeyinde fosforilasyon ve bileşik piruvik asit ( veya tuz formu, piruvat ) ile ikinci bir ATP molekülünün üretilmesiyle sonuçlanır. Enzimatik yollardaki birçok enzim, ters reaksiyonlar için adlandırılır, çünkü enzim hem ileri hem de geri reaksiyonları katalize edebilir.
Glikolizin Sonuçları
Glikoliz glikoz ile başlar ve iki piruvat molekülü, toplam dört ATP molekülü ve iki NADH molekülü ile sona erer. Altı karbonlu halkayı bölünmeye hazırlamak için yolun ilk yarısında iki ATP molekülü kullanıldı, bu nedenle hücrenin kullanımı için iki ATP molekülü ve iki NADH molekülü net kazancı var.
Hücre, piruvat moleküllerini daha fazla katabolize edemezse, bir glikoz molekülünden sadece iki ATP molekülü toplayacaktır. Olgun memeli kırmızı kan hücreleri, organizmaların oksijen varlığında enerjiyi dönüştürdüğü süreç olan aerobik solunum yeteneğine sahip değildir ve glikoliz, tek ATP kaynağıdır. Glikoliz kesintiye uğrarsa, bu hücreler sodyum-potasyum pompalarını koruma yeteneklerini kaybederler ve sonunda ölürler.
Glikolizdeki son adım, piruvat oluşumunu katalize eden enzim olan piruvat kinaz yeterli miktarda mevcut değilse gerçekleşmeyecektir. Bu durumda, tüm glikoliz yolu ilerleyecek, ancak ikinci yarıda sadece iki ATP molekülü yapılacaktır. Bu nedenle, piruvat kinaz, glikoliz için hız sınırlayıcı bir enzimdir.
ÖZETLE: Glikoliz
Glikoliz, enerji elde etmek için glikozun parçalanmasında kullanılan ilk yoldur. Muhtemelen evrimleşen en eski metabolik yollardan biriydi ve dünyadaki neredeyse tüm organizmalar tarafından kullanılıyordu. Glikoliz iki bölümden oluşur: İlk bölüm, altı karbonlu glikoz halkasını iki üç karbonlu şekere bölünme için hazırlar. ATP, ayrılığa enerji vermek için bu yarıda sürece yatırılır. Glikolizin ikinci yarısı, hidrojen atomlarından ATP ve yüksek enerjili elektronları çıkarır ve bunları NAD+'ye bağlar. İlk yarıda iki ATP molekülü yatırılır ve ikinci yarıda substrat fosforilasyonu ile dört ATP molekülü oluşturulur. Bu, hücre için iki ATP ve iki NADH molekülünün net kazancını üretir.
Şekil 4, tüm glikoliz sürecini tek bir görüntüde göstermektedir:

Piruvat Oksidasyonu
Oksijen mevcutsa, aerobik solunum ilerleyecektir. Ökaryotik hücrelerde, glikoliz sonunda üretilen piruvat molekülleri, hücresel solunum bölgeleri olan mitokondriye taşınır. Orada piruvat, koenzim A ( CoA ) adı verilen bir taşıyıcı bileşik tarafından alınacak ve aktive edilecek bir asetil grubuna dönüştürülecektir. Elde edilen bileşiğe asetil CoA denir. CoA, B5 vitamini, pantotenik asitten yapılır. Asetil CoA, hücre tarafından çeşitli şekillerde kullanılabilir, ancak ana işlevi, piruvattan türetilen asetil grubunu glikoz katabolizmasında yolun bir sonraki aşamasına iletmektir.
Pirüvatın Parçalanması
Piruvatın ( glikolizin ürünü olan ) Sitrik Asit Döngüsüne ( hücresel solunumdaki bir sonraki yol ) girmesi için birkaç değişikliğe uğraması gerekir. Dönüştürme üç aşamalı bir işlemdir ( Şekil 5 ).

Aşama 1. Pirüvattan bir karboksil grubu çıkarılır ve çevredeki ortama bir karbondioksit molekülü salınır. Bu adımın sonucu, enzime ( piruvat dehidrojenaz ) bağlı iki karbonlu bir hidroksietil grubudur. Bu, orijinal glikoz molekülünden çıkarılacak altı karbondan ilkidir. Bu adım, metabolize edilen her glikoz molekülü için iki kez ilerler ( unutmayın: glikolizin sonunda üretilen iki piruvat molekülü vardır ); Böylece, her iki adımın sonunda altı karbondan ikisi çıkarılmış olacaktır.
Adım 2. NAD+, NADH'ye indirgenmiştir. Hidroksietil grubu bir asetil grubuna oksitlenir ve elektronlar NAD tarafından alınır NADH+ oluşturur. NADH'den gelen yüksek enerjili elektronlar daha sonra ATP üretmek için kullanılacaktır.
Aşama 3. Bir asetil grubu, asetil CoA ile sonuçlanan konenzim A'ya aktarılır. Enzime bağlı asetil grubu, bir asetil CoA molekülü üreterek CoA'ya aktarılır.
Glikoz metabolizmasının ikinci aşamasında, bir karbon atomu çıkarıldığında, hücresel solunumun ana son ürünlerinden biri olan karbondioksit üreten iki oksijen atomuna bağlandığını unutmayın.
Asetil CoA ila CO2
Oksijen varlığında, asetil CoA, asetil grubunu, üç karboksil grubuna sahip altı karbonlu bir molekül olan sitrat oluşturmak üzere dört karbonlu bir molekül olan oksaloasetata iletir; Bu yol, çıkarılabilir enerjinin geri kalanını bir glikoz molekülü olarak başlayan şeyden toplayacaktır. Bu tek yol farklı isimlerle anılır, ancak biz buna öncelikle Sitrik Asit Döngüsü diyeceğiz.
ÖZETLE: Pirüvat Oksidasyonu
Oksijen varlığında piruvat, bir taşıyıcı koenzim A molekülüne bağlı bir asetil grubuna dönüştürülür. Elde edilen asetil CoA birkaç yola girebilir, ancak çoğu zaman asetil grubu daha fazla katabolizma için sitrik asit döngüsüne iletilir. Pirüvatın asetil grubuna dönüştürülmesi sırasında, bir karbondioksit molekülü ve iki yüksek enerjili elektron çıkarılır. Karbondioksit, orijinal glikoz molekülünün altı karbonundan ikisini ( iki piruvat molekülünün dönüşümü ) oluşturur. Elektronlar NAD+ tarafından alınır ve NADH, elektronları ATP üretimi için daha sonraki bir yola taşır. Bu noktada, başlangıçta hücresel solunuma giren glikoz molekülü tamamen oksitlenmiştir. Glikoz molekülü içinde depolanan kimyasal potansiyel enerji, elektron taşıyıcılarına aktarılmış veya birkaç ATP'yi sentezlemek için kullanılmıştır.
Sitrik Asit Döngüsü
Piruvatın asetil CoA'ya dönüşümü gibi, sitrik asit döngüsü de mitokondri matrisinde gerçekleşir. Bu tek yol farklı isimlerle adlandırılır: sitrik asit döngüsü ( asetat oksaloasetata katıldığında oluşan ilk ara ürün için sitrik asit veya sitrat ), TCA döngüsü ( sitrik asit veya sitrat ve izositrat trikarboksilik asitler olduğundan ) ve Krebs döngüsü, 1930'larda güvercin uçuş kaslarında yoldaki adımları ilk kez tanımlayan Hans Krebs'ten sonra.
Sitrik asit döngüsünün hemen hemen tüm enzimleri, mitokondrinin iç zarına gömülü olan süksinat dehidrojenaz enzimi hariç, çözünür. Glikolizden farklı olarak, sitrik asit döngüsü kapalı bir döngüdür: Yolun son kısmı, ilk adımda kullanılan bileşiği yeniler. Döngünün sekiz adımı, iki karbondioksit molekülü, bir GTP / ATP ve indirgenmiş NADH ve FADH2 formları üreten bir dizi redoks, dehidrasyon, hidrasyon ve dekarboksilasyon reaksiyonudur ( Şekil 6 ). Bu aerobik bir yol olarak kabul edilir, çünkü üretilen NADH ve FADH2, elektronlarını sistemdeki oksijeni kullanacak bir sonraki yola aktarmalıdır. Bu transfer gerçekleşmezse, sitrik asit döngüsünün oksidasyon adımları da gerçekleşmez. Sitrik asit döngüsünün doğrudan çok az ATP ürettiğini ve doğrudan oksijen tüketmediğini unutmayın.

Sitrik Asit Döngüsündeki Adımlar
Aşama 1. İlk adımın başlamasından önce, piruvat oksidasyonu meydana gelmelidir. Ardından, döngünün ilk adımı başlar: Bu, altı karbonlu bir sitrat molekülü oluşturmak için iki karbonlu asetil grubunu dört karbonlu bir oksaloasetat molekülü ile birleştiren bir yoğunlaşma adımıdır. CoA, bir sülfhidril grubuna ( -SH ) bağlanır ve sonunda başka bir asetil grubu ile birleşmek üzere yayılır. Bu adım geri döndürülemez çünkü oldukça eksergoniktir. Bu reaksiyonun hızı, negatif geri besleme ve mevcut ATP miktarı ile kontrol edilir. ATP seviyeleri artarsa, bu reaksiyonun hızı azalır. ATP yetersizse, oran artar.
Adım 2. İkinci adımda, sitrat bir su molekülünü kaybeder ve sitrat izomeri olan izositrata dönüştürülürken bir tane daha kazanır.
Aşama 3. Üçüncü adımda, izositrat oksitlenir ve bir CO2 molekülü ile birlikte beş karbonlu bir molekül olan α-ketoglutarat üretilir ve bu da NAD+'yı NADH'ye indirger. Bu adım aynı zamanda ATP ve NADH'den gelen olumsuz geri bildirimler ve ADP'nin olumlu bir etkisi ile düzenlenir.
Adım 3 ve 4. Üçüncü ve dördüncü adımlar, NAD+'yı NADH'ye indirgeyen elektronları serbest bırakan ve CO2 molekülleri oluşturan karboksil gruplarını serbest bırakan hem oksidasyon hem de dekarboksilasyon adımlarıdır.. α-Ketoglutarat, üçüncü adımın ürünüdür ve bir süksinil grubu, dördüncü adımın ürünüdür. CoA, süksinil CoA'yı oluşturmak için süksinil grubunu bağlar. Dördüncü adımı katalize eden enzim, ATP, süksinil CoA ve NADH'nin geri besleme inhibisyonu ile düzenlenir.
Adım 5. Beşinci adımda, koenzim A yerine bir fosfat grubu ikame edilir ve yüksek enerjili bir bağ oluşturulur. Bu enerji, guanin trifosfat ( GTP ) veya ATP oluşturmak için substrat düzeyinde fosforilasyonda ( süksinil grubunun süksinata dönüştürülmesi sırasında ) kullanılır. Bu adım için, içinde bulundukları hayvan dokusunun türüne bağlı olarak, izoenzimler adı verilen enzimin iki formu vardır. Bir form, kalp ve iskelet kası gibi büyük miktarlarda ATP kullanan dokularda bulunur. Bu form ATP üretir. Enzimin ikinci formu, karaciğer gibi çok sayıda anabolik yola sahip dokularda bulunur. Bu form GTP üretir. GTP, enerjik olarak ATP'ye eşdeğerdir; ancak kullanımı daha kısıtlıdır. Özellikle, protein sentezi öncelikle GTP'yi kullanır.
Adım 6. Altıncı adım, süksinatı fumarata dönüştüren bir dehidrasyon işlemidir. İki hidrojen atomu FAD'ye aktarılarak FADH2 üretilir. Bu atomların elektronlarında bulunan enerji, NAD+ 'yı azaltmak için yetersizdir, ancak FAD'yi azaltmak için yeterlidir. NADH'den farklı olarak, bu taşıyıcı enzime bağlı kalır ve elektronları doğrudan elektron taşıma zincirine aktarır. Bu işlem, mitokondrinin iç zarı içinde bu adımı katalize eden enzimin lokalizasyonu ile mümkün olur.
Adım 7. Yedinci adımda fumarata su eklenir ve malat üretilir. Sitrik asit döngüsündeki son adım, malatı oksitleyerek oksaloasetatı yeniler. İşlemde başka bir NADH molekülü üretilir.
Sitrik Asit Döngüsünün Ürünleri
Her asetil grubundan sitrik asit döngüsüne iki karbon atomu girer ve bir glikoz molekülünün altı karbonundan dördünü temsil eder. Döngünün her dönüşünde iki karbondioksit molekülü salınır; Bununla birlikte, bunlar mutlaka en son eklenen karbon atomlarını içermez. İki asetil karbon atomu, döngünün sonraki dönüşlerinde sonunda serbest bırakılacaktır; Böylece, orijinal glikoz molekülünden gelen altı karbon atomunun tümü sonunda karbondioksite dahil edilir. Döngünün her dönüşü üç NADH molekülü ve bir FADH2 molekülü oluşturur. Bu taşıyıcılar, ATP molekülleri üretmek için aerobik solunumun son kısmı ile bağlantı kuracaktır. Her döngüde bir GTP veya ATP de yapılır. Sitrik asit döngüsündeki ara bileşiklerin birçoğu, esansiyel olmayan amino asitlerin sentezlenmesinde kullanılabilir; Bu nedenle, döngü amfiboliktir ( hem katabolik hem de anabolik ).
ÖZETLE: Sitrik Asit Döngüsü
Sitrik asit döngüsü, yüksek enerjili elektronları ve karbondioksiti uzaklaştıran bir dizi redoks ve dekarboksilasyon reaksiyonudur. NADH ve FADH2 moleküllerinde geçici olarak depolanan elektronlar, sonraki bir yolda ATP üretmek için kullanılır. GTP veya ATP'nin bir molekülü, döngünün her dönüşünde substrat düzeyinde fosforilasyon ile üretilir. Döngüsel yolun doğrusal olanla karşılaştırılması yoktur.
Elektron Taşıma Zinciri
Hücresel solunumda ATP üreten iki yol ( glikoliz ve sitrik asit döngüsü ) hakkında bir şeyler okudunuz. Bununla birlikte, glikozun aerobik katabolizması sırasında üretilen ATP'nin çoğu doğrudan bu yollardan üretilmez. Daha ziyade, elektronları redoks reaksiyonlarına giren bir dizi elektron taşıyıcısı boyunca hareket ettirmekle başlayan bir süreçten türetilir: elektron taşıma zinciri. Bu, hidrojen iyonlarının matris uzayı içinde birikmesine neden olur. Bu nedenle, hidrojen iyonlarının ATP sentazdan geçerek matris boşluğundan yayıldığı bir konsantrasyon gradyanı oluşur. Hidrojen iyonlarının akımı, ADP'yi fosforile eden ve ATP üreten ATP sentazın katalitik etkisine güç sağlar.
Elektron Taşıma Zinciri

Elektron taşıma zinciri ( Şekil 7 ) aerobik solunumun son bileşenidir ve atmosferik oksijen kullanan glikoz metabolizmasının tek parçasıdır. Oksijen sürekli olarak bitkilere yayılır; Hayvanlarda vücuda solunum sistemi yoluyla girer. Elektron taşınması, elektronların bir bileşenden diğerine, elektronların moleküler oksijeni indirgeyerek su ürettiği zincirin uç noktasına hızla geçtiği bir röle yarışına veya kova tugayına benzeyen bir dizi redoks reaksiyonudur. Şekil 7'de I'den IV'e kadar etiketlenmiş proteinlerden oluşan dört kompleks vardır ve bu dört kompleksin bir araya gelmesi, ilişkili hareketli, aksesuar elektron taşıyıcıları ile birlikte elektron taşıma zinciri olarak adlandırılır. Elektron taşıma zinciri, ökaryotların iç mitokondriyal zarında ve prokaryotların plazma zarında birden fazla kopya halinde bulunur. Bununla birlikte, prokaryotların elektron taşıma zincirinin, bazıları anaerobik koşullarda yaşadığı için oksijene ihtiyaç duymayabileceğini unutmayın. Tüm elektron taşıma zincirlerinin ortak özelliği, bir zar boyunca bir proton gradyanı oluşturmak için bir proton pompasının varlığıdır.
Kompleks I
Başlamak için, iki elektron NADH'deki ilk komplekse taşınır. I etiketli bu kompleks, flavin mononükleotidi ( FMN ) ve demir-kükürt ( Fe-S ) içeren bir proteinden oluşur. Riboflavin olarak da adlandırılan B2 vitamininden türetilen FMN, elektron taşıma zincirindeki birkaç protez grubundan veya kofaktörden biridir. Protez grubu, bir proteinin aktivitesi için gerekli olan protein olmayan bir moleküldür. Protez grupları, işlevini kolaylaştıran bir proteine bağlı organik veya inorganik, peptit olmayan moleküllerdir; Protez grupları, enzimlerin protez grupları olan ko-enzimleri içerir. Kompleks I'deki enzim NADH dehidrojenazdır ve 45 amino asit zinciri içeren çok büyük bir proteindir. Kompleks I, matristen zarlar arası boşluğa zar boyunca dört hidrojen iyonu pompalayabilir ve bu şekilde hidrojen iyonu gradyanı, iç mitokondriyal zar tarafından ayrılan iki bölme arasında kurulur ve korunur.
Q ve Kompleks II
Kompleks II, kompleks I'den geçmeyen FADH2'yi doğrudan alır. Birinci ve ikinci kompleksleri üçüncüye bağlayan bileşik ubikinondur ( Q ). Q molekülü yağda çözünür ve zarın hidrofobik çekirdeği boyunca serbestçe hareket eder. İndirgendikten sonra ( QH2 ), ubikinon elektronlarını elektron taşıma zincirindeki bir sonraki komplekse iletir. Q, süksinat dehidrojenaz dahil olmak üzere kompleks I'den NADH'den türetilen elektronları ve kompleks II'den FADH2'den türetilen elektronları alır. Bu enzim ve FADH2, ilk kompleksi atlayarak elektronları doğrudan elektron taşıma zincirine ileten küçük bir kompleks oluşturur. Bu elektronlar baypas ettiğinden ve böylece ilk komplekste proton pompasına enerji vermediğinden, FADH2 elektronlarından daha az ATP molekülü yapılır. Sonuçta elde edilen ATP moleküllerinin sayısı, iç mitokondriyal zar boyunca pompalanan proton sayısı ile doğru orantılıdır.
Kompleks III
Üçüncü kompleks, sitokrom b, başka bir Fe-S proteini, Rieske merkezi ( 2Fe-2S merkezi ) ve sitokrom c proteinlerinden oluşur; Bu kompleks aynı zamanda sitokrom oksidoredüktaz olarak da adlandırılır. Sitokrom proteinleri, protez bir heme grubuna sahiptir. Hem molekülü, hemoglobindeki heme'ye benzer, ancak oksijen değil elektron taşır. Sonuç olarak, çekirdeğindeki demir iyonu elektronları geçerken indirgenir ve oksitlenir, farklı oksidasyon durumları arasında dalgalanır: Fe++ ( indirgenmiş ) ve Fe+++ ( oksitlenmiş ). Sitokromlardaki hem molekülleri, onları bağlayan farklı proteinlerin etkilerinden dolayı biraz farklı özelliklere sahiptir ve her komplekse biraz farklı özellikler verir. Kompleks III, protonları zardan pompalar ve elektronlarını dördüncü protein ve enzim kompleksine taşınmak üzere sitokrom c'ye geçirir ( sitokrom c, Q'dan elektronların alıcısıdır; ancak, Q elektron çiftleri taşırken, sitokrom c bir seferde yalnızca birini kabul edebilir ).
Kompleks IV
Dördüncü kompleks, sitokrom proteinleri c, a ve a3'ten oluşur. Bu kompleks iki hem grubu ( iki sitokromun her birinde bir tane, a ve a3 ) ve üç bakır iyonu ( sitokrom a3 içinde bir çift CuA ve bir CuB ) içerir. Sitokromlar, oksijen tamamen azalana kadar demir ve bakır iyonları arasında bir oksijen molekülünü çok sıkı tutar. İndirgenmiş oksijen daha sonra su ( H2O ) yapmak için çevredeki ortamdan iki hidrojen iyonu alır. Hidrojen iyonlarının sistemden uzaklaştırılması, kemiozmoz sürecinde kullanılan iyon gradyanına katkıda bulunur.
Kemiozmoz
Kemiozmozda, az önce tarif edilen redoks reaksiyonları serisinden elde edilen serbest enerji, membran boyunca hidrojen iyonlarını ( protonlar ) pompalamak için kullanılır. H+ iyonlarının zar boyunca eşit olmayan dağılımı, hidrojen iyonlarının pozitif yükü ve zarın bir tarafında toplanması nedeniyle hem konsantrasyon hem de elektriksel gradyanlar ( dolayısıyla bir elektrokimyasal gradyan ) oluşturur.
Zar, hidrojen iyonları tarafından difüzyona açık olsaydı, iyonlar, elektrokimyasal gradyanları tarafından yönlendirilen matrise geri yayılma eğiliminde olurdu. Birçok iyonun, iyon kanallarının yardımı olmadan fosfolipid membranların polar olmayan bölgelerinden yayılamayacağını hatırlayın. Benzer şekilde, matriks uzayındaki hidrojen iyonları, ATP sentaz adı verilen integral bir membran proteini aracılığıyla yalnızca iç mitokondriyal membrandan geçebilir ( Şekil 8 ). Bu karmaşık protein, içinden yayılan hidrojen iyonlarının kuvvetiyle elektrokimyasal gradyanlarından aşağı doğru döndürülen küçük bir jeneratör görevi görür. Bu moleküler makinenin parçalarının döndürülmesi, hidrojen iyonu gradyanının potansiyel enerjisini kullanarak ATP'yi oluşturan ADP'ye bir fosfat eklenmesini kolaylaştırır.
Alıştırma Sorusu

Soru
Dinitrofenol ( DNP ), iç mitokondriyal zarın protonlara sızdırmasını sağlayan bir bağlayıcıdır. 1938 yılına kadar zayıflama ilacı olarak kullanıldı. DNP'nin iç mitokondriyal zar boyunca pH'daki değişim üzerinde nasıl bir etkisi olmasını beklersiniz? Bunun neden etkili bir kilo verme ilacı olabileceğini düşünüyorsunuz?
Cevabı göster
DNP zehirlenmesinden sonra, elektron taşıma zinciri artık bir proton gradyanı oluşturamaz ve ATP sentaz artık ATP yapamaz. DNP etkili bir diyet ilacıdır çünkü ATP sentezini ayrıştırır; Başka bir deyişle, aldıktan sonra, kişi yediği yiyeceklerden daha az enerji elde eder. İlginç bir şekilde, bu ilacın en kötü yan etkilerinden biri hipertermi veya vücudun aşırı ısınmasıdır. ATP oluşturulamadığından, elektron taşınımından gelen enerji ısı olarak kaybolur.
Kemiozmoz ( Şekil 9 ), aerobik glikoz katabolizması sırasında yapılan ATP'nin yüzde 90'ını üretmek için kullanılır; Aynı zamanda, fotofosforilasyon sürecinde güneş ışığının enerjisinden yararlanmak için fotosentezin ışık reaksiyonlarında kullanılan yöntemdir. Mitokondride kemiozmoz sürecini kullanarak ATP üretiminin oksidatif fosforilasyon olarak adlandırıldığını hatırlayın. Bu reaksiyonların genel sonucu, hidrojen atomlarından çıkarılan elektronların enerjisinden ATP üretimidir. Bu atomlar başlangıçta bir glikoz molekülünün parçasıydı. Yolun sonunda, elektronlar bir oksijen molekülünü oksijen iyonlarına indirgemek için kullanılır. Oksijen üzerindeki ekstra elektronlar, çevredeki ortamdan hidrojen iyonlarını ( protonları ) çeker ve su oluşur.
Alıştırma Sorusu

Soru
Siyanür, elektron taşıma zincirinin bir bileşeni olan sitokrom c oksidazı inhibe eder. Siyanür zehirlenmesi meydana gelirse, zarlar arası boşluğun pH'ının artmasını veya azalmasını bekler misiniz? Siyanürün ATP sentezi üzerindeki etkisi nedir?
Cevabı göster
Siyanür zehirlenmesinden sonra, elektron taşıma zinciri artık elektronları zarlar arası boşluğa pompalayamaz. Zarlar arası boşluğun pH'ı artacak, pH gradyanı azalacak ve ATP sentezi duracaktır.
ATP Verimi
Glikozun katabolizmasından üretilen ATP moleküllerinin sayısı değişir. Örneğin, elektron taşıma zinciri komplekslerinin zardan pompalayabileceği hidrojen iyonlarının sayısı türler arasında değişir. Başka bir varyans kaynağı, mitokondri zarları boyunca elektronların mekiğinden kaynaklanmaktadır. ( Glikolizden üretilen NADH mitokondriye kolayca giremez. ) Böylece, elektronlar mitokondrinin içinde NAD+ veya FAD+ tarafından alınır. Daha önce öğrendiğiniz gibi, bu FAD+ molekülleri daha az iyon taşıyabilir; sonuç olarak, FAD+ taşıyıcı olarak hareket ettiğinde daha az ATP molekülü üretilir. NAD+ karaciğerde elektron taşıyıcısı olarak kullanılır ve FAD+ beyinde etki eder.
Glikozdan üretilen ATP moleküllerinin verimini etkileyen bir diğer faktör, bu yollardaki ara bileşiklerin başka amaçlar için kullanılmasıdır. Glikoz katabolizması, hücrelerdeki diğer tüm biyokimyasal bileşikleri oluşturan veya parçalayan yollarla bağlantı kurar ve sonuç, şimdiye kadar açıklanan ideal durumlardan biraz daha karmaşıktır. Örneğin, glikoz dışındaki şekerler, enerji ekstraksiyonu için glikolitik yola beslenir. Ayrıca, nükleik asitleri oluşturan beş karbonlu şekerler, glikolizdeki ara maddelerden yapılır. Bazı esansiyel olmayan amino asitler, hem glikoliz hem de sitrik asit döngüsünün ara ürünlerinden yapılabilir. Kolesterol ve trigliseritler gibi lipitler de bu yollardaki ara ürünlerden yapılır ve hem amino asitler hem de trigliseritler bu yollardan enerji için parçalanır. Genel olarak, canlı sistemlerde, bu glikoz katabolizma yolları, glikozda bulunan enerjinin yaklaşık yüzde 34'ünü çıkarır.
ÖZETLE: Elektron Taşıma Zinciri
Elektron taşıma zinciri, glikoz katabolizmasında ara bileşiklerden çıkarılan elektronların son elektron alıcısı olarak serbest oksijeni kullanan aerobik solunumun bir kısmıdır. Elektron taşıma zinciri, iç mitokondriyal zara gömülü dört büyük, multiprotein kompleksinden ve aralarında elektron taşıyan iki küçük yayılabilir elektron taşıyıcısından oluşur. Elektronlar, hidrojen iyonlarını bir zar boyunca taşımak için üç noktada kullanılan az miktarda serbest enerji ile bir dizi redoks reaksiyonundan geçirilir. Bu işlem, kemiozmozda kullanılan gradyana katkıda bulunur. Elektron taşıma zincirinden geçen elektronlar yavaş yavaş enerji kaybeder, NADH veya FADH2 tarafından zincire bağışlanan yüksek enerjili elektronlar zinciri tamamlar, çünkü düşük enerjili elektronlar oksijen moleküllerini azaltır ve su oluşturur. Elektronların serbest enerji seviyesi, NADH'de yaklaşık 60 kcal/mol'den veya FADH2 45'de kcal/mol'den suda yaklaşık 0 kcal/mol'e düşer. Elektron taşıma zincirinin son ürünleri su ve ATP'dir. Sitrik asit döngüsünün bir dizi ara bileşiği, esansiyel olmayan amino asitler, şekerler ve lipitler gibi diğer biyokimyasal moleküllerin anabolizmasına yönlendirilebilir. Aynı moleküller, glikoz yolları için enerji kaynakları olarak hizmet edebilir.
Hadi İnceleyelim
Hücresel solunum, üç benzersiz metabolik yoldan oluşan bir koleksiyondur: glikoliz, sitrik asit döngüsü ve elektron taşıma zinciri. Glikoliz anaerobik bir süreçtir, diğer iki yol ise aerobiktir. Glikolizden sitrik asit döngüsüne geçmek için, piruvat moleküllerinin ( glikoliz çıkışı ) piruvat oksidasyonu adı verilen bir işlemle oksitlenmesi gerekir.
Glikoliz
Glikoliz, hücresel solunumdaki ilk yoldur. Bu yol anaerobiktir ve hücrenin sitoplazmasında gerçekleşir. Bu yol 1 glikoz molekülünü parçalar ve 2 piruvat molekülü üretir. Her yarıda beş adım bulunan iki yarım glikoliz vardır. İlk yarı "enerji gerektiren" adımlar olarak bilinir. Bu yarı glikozu ayırır ve 2 ATP kullanır. Piruvat kinaz konsantrasyonu yeterince yüksekse, glikolizin ikinci yarısı devam edebilir. İkinci yarıda, "enerji salınımı: adımlar, 4 ATP molekülü ve 2 NADH salınır. Glikoliz, 2 ATP molekülü ve 2 NADH net kazancına sahiptir.
Bazı hücreler ( örneğin, olgun memeli kırmızı kan hücreleri ) aerobik solunum yapamazlar, bu nedenle glikoliz onların tek ATP kaynağıdır. Bununla birlikte, çoğu hücre piruvat oksidasyonuna uğrar ve hücresel solunumun diğer yollarına devam eder.
Piruvat Oksidasyonu
Ökaryotlarda mitokondride piruvat oksidasyonu gerçekleşir. Piruvat oksidasyonu ancak oksijen mevcutsa gerçekleşebilir. Bu işlemde, glikoliz tarafından oluşturulan piruvat oksitlenir. Bu oksidasyon işleminde, piruvattan bir karboksil grubu çıkarılır ve asetil CoA oluşturmak üzere koenzim A ( CoA ) ile birleşen asetil grupları oluşturulur. Bu işlem aynı zamanda CO2'yi de serbest bırakır.
Sitrik Asit Döngüsü
Sitrik asit döngüsü ( Krebs döngüsü olarak da bilinir ) hücresel solunumdaki ikinci yoldur ve aynı zamanda mitokondride de gerçekleşir. Döngünün hızı ATP konsantrasyonu ile kontrol edilir. Daha fazla ATP mevcut olduğunda, hız yavaşlar; daha az ATP olduğunda oran artar. Bu yol kapalı bir döngüdür: son adım, ilk adım için gereken bileşiği üretir.
Sitrik asit döngüsü aerobik bir yol olarak kabul edilir, çünkü ürettiği NADH ve FADH2, elektronlarını atmosferik oksijen kullanan bir sonraki yola ( elektron taşıma zinciri ) aktaran geçici elektron depolama bileşikleri olarak işlev görür. Sitrik asit döngüsünün her dönüşü net bir CO2, 1 GTP veya ATP ve 3 NADH ve 1 FADH2 kazancı sağlar.
Elektron Taşıma Zinciri
Glikozdan gelen ATP'nin çoğu elektron taşıma zincirinde üretilir. Hücresel solunumun doğrudan oksijen tüketen tek kısmıdır; Bununla birlikte, bazı prokaryotlarda bu anaerobik bir yoldur. Ökaryotlarda bu yol iç mitokondriyal zarda gerçekleşir. Prokaryotlarda plazma zarında oluşur.
Elektron taşıma zinciri, zar boyunca 4 proteinden ve bir proton pompasından oluşur. Bir kofaktör, elektronları I - III proteinleri arasında taşır. NAD tükenirse, I'i atlayın: FADH2, II'de başlar. Kemiozmozda, bir proton pompası hidrojenleri mitokondrinin içinden dışarıya alır; Bu, "motoru" döndürür ve fosfat grupları buna bağlanır. Hareket, ADP'den ATP'ye değişir ve aerobik glikoz katabolizmasından elde edilen ATP'nin % 90'ını oluşturur.








")





















