Bitkilerde Üreme Nasıl Gerçekleşir Ve Çeşitleri Nelerdir?
Bitkilerde Üreme Nasıl Gerçekleşir Ve Çeşitleri Nelerdir?
- BİLİM ve TEKNOLOJİ
- Mon, 8 Apr 2024 16:04:17
- Mon, 8 Apr 2024 16:04:17
Bitki üremesi, eşeyli veya eşeysiz olarak gerçekleşen bitki yavrularının üretimidir. Eşeysiz üreme, orijinal bitkinin genetik bir klonunu üretirken, eşeyli üreme, gametlerin füzyonu ile gerçekleşir ve ana bitkilerden genetik olarak benzersiz yavrular üretir. Eşeyli üreme, alelleri bir nesilden diğerine yeniden birleştirerek popülasyonun genetik çeşitliliğini arttırır. Eşeyli üreme yoluyla bir popülasyonun genetik çeşitliliğinin arttırılması, bir bütün olarak bitki popülasyonunun, biyotik ( yani hastalıklar ) veya abiyotik ( yani kuraklık gibi lokalize bir rahatsızlık veya iklim değişikliği gibi daha uzun vadeli etkiler ) gibi değişen çevresel koşullara uyum sağlamasına izin verir. Bununla birlikte, eşeyli üreme, gametlerin bir bitkiden diğerine başarılı bir şekilde aktarılmasını gerektirir, bu da bitkilerin hareketsizliği göz önüne alındığında zordur. Eşeyli üreme risklerini en aza indirmek için, birçok bitki eşeyli veya eşeysiz olarak üreme yeteneğine sahiptir. Eşeysiz üreme, bitkilerin döllenme olmadığında üremelerine izin verir, bu da onların lokalize genişlemeyi hızlandırmalarına izin verir. Eşeysiz üremenin en büyük riski, üretilen yeni oluşan bitkilerin genetik olarak ebeveynle aynı olmasıdır ( mutasyonlar hariç ). Bu kısa vadede bir avantaj olsa da, çevresel koşullar sürekli değişiyor ve eşeysiz üremeye büyük ölçüde bağımlı olması nedeniyle düşük genetik çeşitliliğe sahip türler, çevre aniden değişirse veya popülasyona yeni bir hastalık girerse çok sayıda ölebilir.

Eşeysiz Üreme
Eşeysiz üreme, kaynak hücrelerde mutasyonlar meydana gelmedikçe, genetik olarak ana bitkiyle özdeş olan yeni bireysel bitkiler üretir. Eşeysiz üreme bitkilerde iki farklı mekanizma ile gerçekleşebilir: vejetatif üreme ve apomoksis. Vejetatif üreme, bitkinin vejetatif bir kısmının ( yani yaprak, gövde veya kök ) ebeveyn bitkiden çıkarıldığı ve ayrı bir birey oluşturduğu eşeysiz üremenin bir örneğidir. Apomoksis, döllenmemiş gametler ( polen veya ovüller ) genetik olarak özdeş sporlara veya tohumlara dönüştüğünde ortaya çıkar.
Vejetatif üreme türleri
Vejetatif üreme, bitkilerde çeşitli mekanizmalarla gerçekleşir. Birçok bitki, bir kez kurulduktan sonra ana bitkiden uzakta yaşayabilen yeni bireysel bitkilerin geliştirilmesinde uzmanlaşmış değiştirilmiş gövdelere sahiptir. Rizomlar, yeraltından geçen ve yeni bir birey üreten ana bitkiden uzaklaşan değiştirilmiş gövdelerdir. İris bitkileri genellikle bu şekilde çoğalarak koloniler ( veya kümeler ) oluşturur. Kavak ağaçları ( Populus cinsi içinde ), ağırlıklı olarak ana bitkiden 30 - 40 m boyunca yer altında seyahat eden rizomlar yoluyla çoğalan son derece büyük koloniler halinde büyür. Utah'taki bir titrek kavak kolonisinin 80.000 yıldan daha eski olduğu tahmin ediliyor ve dünyanın en büyük organizmaları olarak kabul ediliyor. Köksaplarla üreyen bitkiler de yangına dayanıklıdır. Bir yangın olayı sırasında toprak çizgisinin hemen altındaki sıcaklıklara büyük ölçüde aracılık edilir ve bu bitkiler tipik olarak bir yangından sonra yeni bitkileri yeniden filizlendirebilir. Stolonlar, vejetatif üremeyi destekleyen modifiye edilmiş gövdelerin bir başka örneğidir. Yerin altına yayılmak yerine, dışkılar ( genellikle "koşucular" olarak bilinir ) ana bitkiden yer üstüne yayılır. Bunun en yaygın örneği çimlerde kullanılan yengeç otudur. Çilekler ayrıca dışkılarla çoğalır. Bitkiler ayrıca kök sistemlerinden vejetatif olarak çoğalabilirler. Kökler ve gövdeler anatomik olarak farklıdır, bu nedenle bir köksap kökten farklıdır. Bazı türlerde ( yani güllerde ), kök sistemi toprak profilinin yüzeyine ulaştığında, sapları ve yaprakları oluşturan maceracı tomurcuklar ortaya çıkar ve ardından yeni bir bitki oluşturur. Bu süreç enayi olarak bilinir. Yeraltı soğanı üreten bazı türler ( yani soğan ve laleler ) yeni soğanlar üreterek vejetatif olarak çoğalabilir. Ampuller, yeraltı yaprakları olan kısa bir gövdeden oluşan son derece uzmanlaşmış yeraltı bitki yapılarıdır. Tipik olarak, bir ampul, bitkinin kış mevsiminde uykuda kalmasına izin veren bir depolama mekanizmasıdır. İlkbahar çıktığında yeraltı yaprakları ortaya çıkar ve fotosentez yapmaya başlar.
Apomiksis Çeşitleri
Apomixis, döllenmemiş embriyonun yeni bir bitkiye dönüşmesidir. Bitkiler arasında birçok apomiksis çeşidi vardır. Apomiksis geçiren hemen hemen tüm bitkiler fakültatiftir ( fakültatif apomiksis ), yani apomiksis sadece başarılı bir şekilde döllenmemiş gametlerde meydana gelir. Bazı vasküler olmayan bitki türlerinde ( yani yosun, ciğer otları ve boynuz otları ) ve birkaç çekirdeksiz vasküler bitkide ( yani eğrelti otları ve kulüp yosunları ), gametofitler ( haploid gamet üreten yapılar ) gibi görünen şeyler aslında sporofitlere benzeyen hücreler üretir ( haploid hücreler: yumurta veya sperm ) ancak ana bitki ile aynı kromozom sayısını ( ploidi seviyesi ) korur. Bu apomiksis süreci apogami olarak bilinir ve genetik olarak özdeş sporlar ( tek hücreli embriyolar ) üretir. Bildiğimiz kadarıyla, apomiksis gymnospermlerde tamamen yoktur. Bununla birlikte, anjiyospermlerde ( çiçekli bitkiler ) apomiksisin çeşitli varyasyonları vardır. Gametofiik apomiksiste, diploid bir embriyo, mayoz bölünme geçirmemiş bir hücreden bir tohuma ( embriyo ve endospermde depolanmış gıdayı içeren çok hücreli bir yapı ) dönüşür. Bazı soğan türleri ( Allium sp. ), çiçeklerin döllenmemiş embriyolarının çiçeklenme döneminde vejetatif yapılara ( ampuller olarak bilinir ) dönüştüğü vejetatif apomiksis geçirir. Bu soğanlar, sapları, kökleri ve yaprakları bulunan küçük bitkilere benzer. Ampuller nemli toprakla temas ettiğinde yeni bir bireye dönüşebilirler.
Eşeyli üreme
Bitkilerde eşeyli üreme, genetik olarak farklı iki bitkiden gametlerin ( mayoz bölünme geçirmiş hücreler ) döllenmesinden, birleşmesinden kaynaklanır. Mayoz bölünme, gametlerin yarısını üreten kromozom sayısını azaltır. Döllenme, farklı bireylerden gametleri yeniden birleştirerek, sporlara veya tohumlara dönüşen bir zigot üretir. Eşeyli üremenin temel amacı, bir sonraki nesilde genlerin yeniden düzenlenmesidir. Bu, popülasyonun genetik çeşitliliğini arttırmaya ve değişen çevresel faktörler karşısında türlerin evrimsel canlılığını arttırmaya hizmet eder.
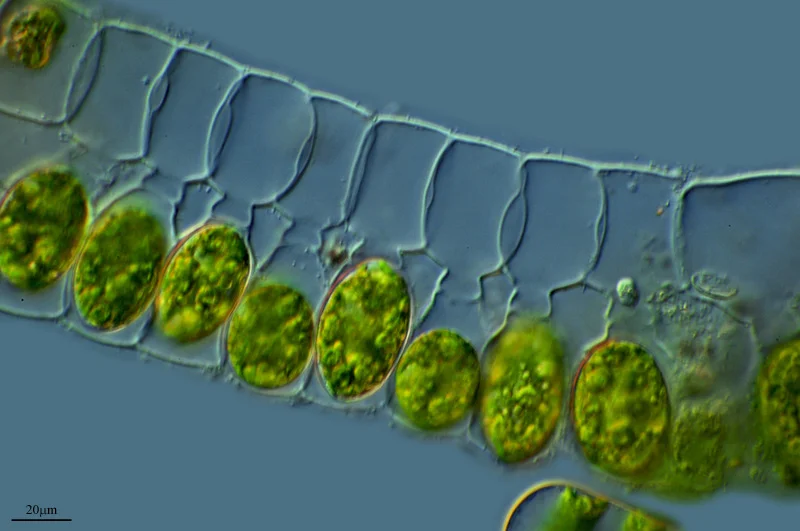
Konjugasyon: yeşil alglerde eşeyli üremenin kökeni
Yeşil algler ( Phylum Chlorophyta ), tek hücreli, kolonyal ( birbirine bağlı aynı türden hücreler ) veya çok hücreli ( bir organizma içindeki farklı hücre tipleri ) olabilen sucul, fotosentetik türlerdir. Yeşil alglerin çoğu eşeysiz üreme yoluyla çoğalırken, bazı gruplar ( özellikle Spirogyra ) eşeyli üreme geliştirdi. Konjugasyon olarak bilinen bir süreçte, bir diploid ( 2n kromozomlu ) yeşil alg hücresi, bir sonraki nesilde genetik olarak benzersiz diploid zigotlar oluşturmak için döllenme yoluyla yeniden birleşen dört haploid ( n kromozom ) hücre üreten mayoz bölünmeye uğrayacaktır. Konjugasyon sırasında, döllenmiş bir diploid ( 2n ) alg hücresi, dikenli bir dış kabuğa sahip bir diploid zigospor üreten hücre duvarını kalınlaştırarak bu işleme başlar. Zigospor daha sonra dikenli hücre duvarının içinde dört haploid ( n ) hücre üreten mayoz bölünmeye uğrar. Bu haploid hücrelerden ikisi bir çiftleşme suşundan ( + ), diğer ikisi ise zıt çiftleşme suşundan ( - ) olacaktır. Döllenme sırasında, + çiftleşme suşları sadece – çiftleşme suşları ile kaynaşabilir. Bu, daha sonra bitkilerde geliştirilen dişi ve erkek gametlerin ilkel bir versiyonudur. Haploid hücreler geliştikten sonra, çimlenme olarak bilinen bir süreçte dikenli hücre duvarı parçalanır ve haploid hücreleri serbest bırakır. Çimlenmeyi takiben, haploid hücreler iki kamçı ile yüzer ve birlikte kümeler oluşturarak toplanır. Her küme, aynı çiftleşme tipindeki haploid hücreleri içerir, + çiftleşme tipi kümeler ve – çiftleşme tipi kümeler oluşturur. Farklı çiftleşme tipi kümeler birbiriyle temas ettiğinde, bir + haploid ve bir – haploid hücre arasında bir hücresel zar tüpü ( sitoplazmik iplik olarak bilinir ) oluşur. Sitoplazmik iplik büyüdükçe, iki hücrenin sitoplazma ve haploid ( n ) çekirdekleri birleşerek döllenme olarak bilinen bir süreçte genetik olarak benzersiz bir diploid ( 2n ) yeşil alg hücresi oluşturur.

Nesillerin Değişimi, çok hücreli yeşil alglerden kaynaklanmaktadır
Konjugasyon, tek hücreli veya kolonyal bir tür içinde bir eşeyli üreme şekli iken, bazı çok hücreli yeşil alg grupları ( ve kara bitkileri ), nesillerin değişimi olarak bilinen bir süreçle çoğalır. Bu süreçte, tek hücreli bir üreme aşaması ile birbirine bağlanan iki çok hücreli nesil vardır. Gametofit olarak bilinen bir nesil, bütün bir haploid ( n ) çok hücreli organizma üretir ve gametangium olarak bilinen özel bir dokuda mitoz yoluyla haploid ( n ) gametler üretir, tam olarak aynı sayıda kromozoma sahip gametofit üreten bir gametofit organizmanın somatik ( veya vücut ) hücreleri. Gametofit neslinden gelen haploid ( n ) gametler, bir diploid ( 2n ) zigot oluşturmak için birleşir. Zigot, sporopihte olarak bilinen bir diploid ( 2n ), çok hücreli organizmaya dönüşürken. Gametofit aşamasından farklı olarak, sporofit, sporangium olarak bilinen özel bir dokuda mayoz bölünme geçirerek, kromozom sayısını yarıya indirerek sporlar üretir. Bu haploid sporlar, haploid gametofit üreterek filizlenir ve döngüyü yeniden başlatır. Çok hücreli yeşil algler ne gametofit dapoominant ne de sporofit baskındır. Kara bitkileri ya gametofit baskın ya da sporofit baskındır.
Stonewortlar çok hücreli üreme yapılarına sahiptir
Stoneworts ( Charophyceae Sınıfı ), yeşil algler ve kara bitkileri arasında gelişimsel bir adımı temsil eden suda yaşayan fotosentetik organizmalardır. Stoneworts, silindirik bir eksenle birbirine bağlı bir düğüm üzerinde ortalanmış filament kıvrımları içeren vejetatif dokulara sahiptir ve bir alt tabakaya bağlanmalarına izin veren tek hücreli, kök benzeri yapılar ( rizoidler ) içerir. Çok hücreli yeşil alglerin aksine, mevcut taş otları nesiller arası değişim yoluyla çoğalmıyor gibi görünmektedir. Aksiyel düğümlere bağlı üreme organları arasında oogonium ve antheridium bulunur. Oogonina ve antheridia'nın her ikisi de steril ( kısır ) hücrelerden oluşan bir zarfla çevrilidir. Oogoniaum, megaspor olarak bilinen nispeten büyük bir hareketsiz yumurta içeren çok hücreli, dişi bir cinsel organdır. Oogonianum, spiral filamentlerden oluşan bir zarftan oluşan büyük vazo şeklinde ( veya oval ) bir yapıdır, antheridium ise daha küçük bir küresel yapıdır. Ayrı erkek cinsiyet organları ( antheridia ) her biri, mikrospor olarak bilinen nispeten küçük bir sperm hücresi içerir. Gametler çok hücreli bir yapı içinde korunur, yeşil alglerde görülmez, bu da Phylum Chlorophyta'ya dahil edilmelerine karşı ana argümandır. Yeşil alglere göre, taş otları son derece büyük yumurtalara sahiptir. Hareketli, sperm hücreleri ( n ) hareketsiz yumurta hücrelerine ( n ) yüzerek onları dölleyerek yeni bir genetik, diploid birey üreterek filizlenebilen bir diploid ( 2n ) zigot oluşturur. Charophyceae içindeki türler ya tek evcikli ( antheridia ve oogonia aynı bitkide bulunur ) ya da ikievcikli ( bireyler ya antheridia ya da oogonia üretir, ancak ikisini birden üretmez ).
Kara bitkilerinin embriyoları besleyici, koruyucu dokularla sarılır
Gerçek bitkiler ( Sınıf Embryophyceae ), karada kalmalarına izin veren çeşitli morfolojilere sahiptir. Üreme açısından, kara bitkileri, bitki embriyolarının kendilerini çevreleyen dokudan besin elde etmeleri ve gelişimlerinin erken aşamalarında hayatta kalma yeteneklerini artırmaları nedeniyle taş otlarından farklıdır. Besleyici dokuya ek olarak, bitki embriyoları, embriyonun karadayken susuz kalmasına yardımcı olan koruyucu bir dış dokuya sahiptir. Mevcut ( hala yaşayan ) taş otları, nesiller arası değişim yoluyla üremiyor gibi görünse de, tüm kara bitkileri çoğalır. Çoğu taksonomist, kara bitkilerinin taş otlarından ortaya çıktığı sonucuna varır ( genetik analizler ve üreme organlarının çok hücreli doğası nedeniyle ), ataların taş otlarının muhtemelen nesiller arası değişim yoluyla çoğaldığını, ancak bu taksonların artık neslinin tükendiğini öne sürer.
Briyofitler sudan karaya geçti
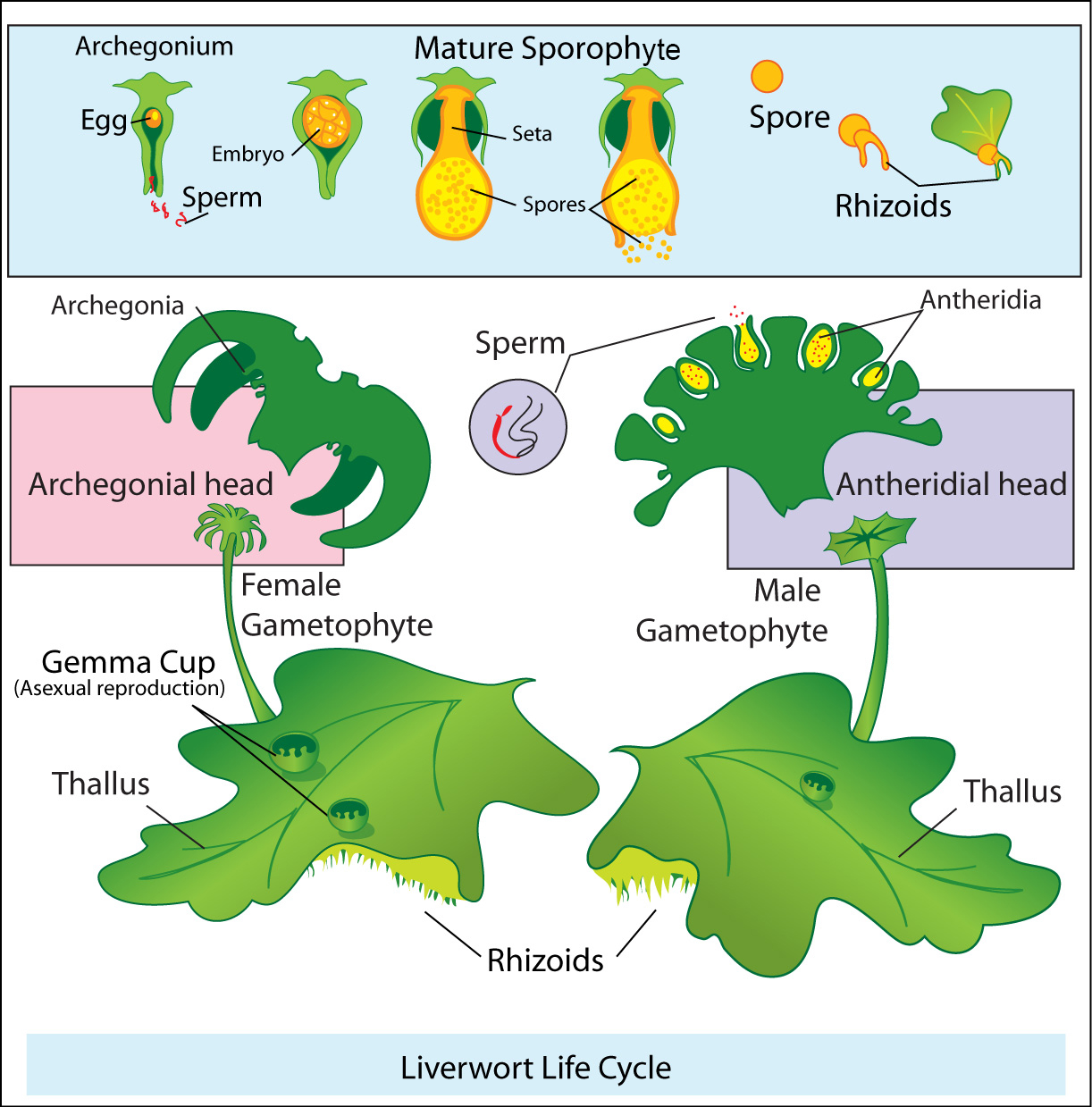
Briyofitler ( veya vasküler olmayan bitkiler ) taş otlarından ortaya çıktı ve ilk gerçek kara bitkilerini temsil ediyor. Briyofitler, gerçek vasküler dokudan yoksun küçük kara bitkileridir, sporlar yoluyla çoğalırlar ve gerçek yosunları ( Phylum Bryophyta ), ciğer otlarını ( Phylum Hepatophyta ) ve boynuz otlarını ( Phylum Anthocerotophyta ) içerir. Karaya geçiş, bitkilerin yeterli bir ozmotik ( H2O ) dengesini korumasına izin veren taş otlarında bulunmayan benzersiz morfolojiler gerektiriyordu. Vejetatif olarak, kara bitkilerinin yaprakların ( ve bazı gövdelerin, yani kaktüslerin ) dış kısmında, suyun bitkiden dışarı yayılmasını önleyen hava geçirmez bir sistem oluşturan mumsu bir kütikül tabakası vardır. Bununla birlikte, bitkilerin fotosentez yapmak için CO2'e ihtiyacı vardır. Koruyucu hücreler olarak bilinen özel hücreler, yaprağın açılıp kapanan epidermisinde bulunur ve yaprakta stoma olarak bilinen gözenekler oluşturur. Bu, CO2'nin içeri girmesine izin verirken, aynı zamanda suyu da serbest bırakır. Bitkiler, mevcut toprak nemi mevcudiyetine bağlı olarak yaprak içindeki ozmotik dengeyi düzenlemek için stomalarını sürekli olarak açıp kapatırlar. Tüm kara bitkileri, nesiller arası dönüşümle çoğalır. Haploid gametofitler, birkaç eşleşmemiş kromozoma sahip hücrelere sahiptir ve mitoz yoluyla haploid gametler üretir. Bu gametler döllenme sırasında birleşir ve diploid somatik hücreler içeren bir sporofit haline gelen bir zigot üretir. Özel dokular mayoz bölünmeye uğrar ve döllenme olmadan çimlenen dört haloid sporu üretir.
Briyofit yaşam döngüsü gametofit baskındır, yani yaşam döngüsünün çoğunluğuna gametofit nesli hakimdir. Gametofitler, sporofit neslinden daha belirgin ve daha uzun ömürlüdür. Sporofitler aralıklı olarak ortaya çıkar ve tipik olarak gelişim için besinsel olarak gametofitlere bağımlıdır. Briyofitlerin gametofitleri ya erkek ya da dişidir. Gametofitlerin fotosentetik dokuları hemen hemen aynı olsa da, üreme organları her cinsiyete özgüdür ve sürgünlerin uçlarında veya yaprakların akslarında bulunur. Erkek gametofitler, birkaç erkek gametten veya steril, kısır bir doku ile kaplanmış spermden oluşan oval yapılar olan antheridia olarak bilinen cinsel organlar üretir. Buna karşılık, bir taş otunun antheridium'u sadece bir erkek gamet üretir. Dişi gametofitler, steril bir doku içinde tek bir yumurta yuvası ile archegonium olarak bilinen vazo şeklinde bir cinsel organ üretir. Briyofitlerin erkek gametleri ( yeşil algler ve taş otlarının yanı sıra ) archegonium'a ulaşmak için suya ihtiyaç duyar. Suyun varlığında, briyofitlerin erkek gametleri, archegonium tarafından üretilen cinsel bir feromon ( kimyasal ) tarafından yönlendirilen antheridium'a doğru yüzer.
Erkek ve dişi gametler döllenme yoluyla birleştikten sonra, sporofit embriyosu olarak bilinen diploid bir zigot üretirler. Bu embriyo archegonium'un içinde gelişir ve sonunda bir sporofit üretir. Sporofitler tipik olarak göze çarpmaz ve fotosentetik değildir. Sonuç olarak, besinsel olarak gametofitlere bağımlıdırlar ve bağlı kalırlar. Sporofit gelişiminin sonunda, iç hücrelerin mayoz bölünmeye maruz kaldığı, erkek ve dişi haploid ( n ) sporları üreten sporangium olarak bilinen çok hücreli bir yapı gelişir. Bu sporlar tipik olarak rüzgarla dağılır ve filizlenir ve briyofit yaşam döngüsünü yeniden başlatan bir gametofit haline gelir.
Likofitlerde megagametofitler ve mikrogametofitler
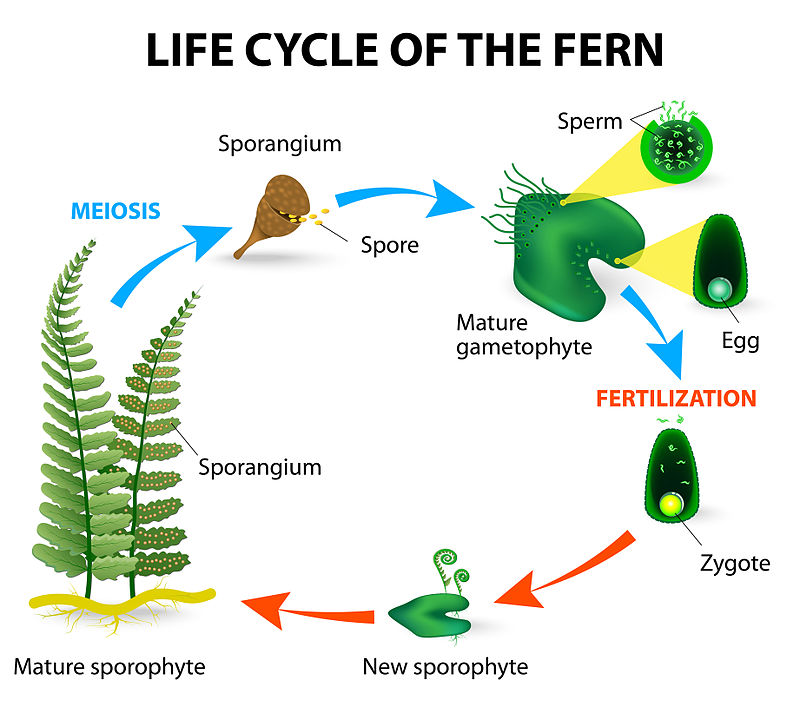
Tüm vasküler olmayan bitkiler ve çekirdeksiz vasküler bitkilerin çoğu homosporludur, bir tür spor ( erkek veya dişi ) üretir ve mitoz yoluyla gametofitin ortaya çıkmasına neden olur. Lycopodiopsida içindeki bazı kulüp yosunu ( yani Selaginella ) cinsleri, ayrı erkek ve dişi gametofitler üreten iki farklı türde spor ( heterospori ) üreten sporofitik bitkilerdir. Heterosporlu likofitlerin gametofiti, sporangium bitkisinde kapalı kalırken oldukça azalır. Sporofit olgunlaştığında, megasporangium olarak bilinen çok hücreli yapılar geliştirecektir. Megasporangiumun içinde, megasporofit olarak bilinen olgunlaşmamış bir yumurta hücresi bulunur. Bu hücre, megasporlar olarak bilinen dört haploid yumurta hücresi üreten mayoz bölünmeye uğrar. Mayoz bölünme meydana geldiğinde, bu yapı bir megagametofit olarak bilinir. Megagametofitler, dört megaspor ( veya yumurta ) üreten ve barındıran büyük çok hücreli yapılardır, mikrogametofitler ise sperm üreten ve barındıran çok daha küçük, çok hücreli yapılardır. Mikrogametofitler, mikrosporangium içindeki mikrosporositlerin mayoz bölünmeye maruz kaldığı ve haploid mikrosporlar ürettiği diploid mikrosporangiden kaynaklanır.
Çekirdeksiz vasküler bitkilerin çoğu ( Lycopodiopsida'nın homosporlu üyeleri dahil ) ayrı gametofitik ve sporofitik formlara sahiptir. Bu formlar fiziksel olarak birbirine bağlı olabilirken, megagametofitler ve mikrogametofitler üreten likofitler, mikro ve megagametofitlerde görülen sporofitik doku içinde bulunan oldukça indirgenmiş gametofitler üretir. Yumurtanın döllenmesi gerçekleştiğinde, arkegonyumun diploid dokusu sporofitik embriyoyu sararak büyümeye devam eder ve olgun bir megagametofit haline gelir. Olgun megagametofit, megaspor duvarı olarak bilinen tek bir epidermal hücre tabakası ile çevrilidir. Döllenmiş megagametofit filizlendikten sonra, gelişmekte olan embriyo, tohumlu bitkilere benzer şekilde megagametofitlerin kısır dokusunda depolanan yiyeceklerle desteklenir. Diğer çekirdeksiz vasküler bitkiler bu besin rezervine sahip değildir, bu da bu likofitlere gelişimin erken aşamalarında diğer çekirdeksiz vasküler bitkilere göre rekabet avantajı sağlar.

Gymnospermlerin çıplak tohumları vardır
Mevcut tohumlu bitkiler ( veya spermatofitler ) arasında açık tohumlular ve kapalı tohumlular bulunur. Fosil kanıtları, tohumlu bitkilerin artık soyu tükenmiş tohumlu eğrelti otlarından kaynaklandığını göstermektedir. Gymnospermler ( Yunanca "çıplak tohumlar" ) en ilkel, mevcut tohumlu bitkilerdir ve kapatılmamış tohumlar üretirken, kapalı tohumlular ( Yunanca "damarlı tohumlar" ) tohumları meyve olarak bilinen ek bir dokuda kaplanmış ( veya kaplı ) olan çiçekli bitkilerdir. Mevcut açık tohumlular arasında sikadlar ( Alt Sınıf Cycadidae ), gingko ağacı ( Alt sınıf Ginkgoidae ), gnetofitler ( Alt sınıf Gnetidae ) ve kozalaklı ağaçlar ( Alt sınıf Pinidae ) bulunur.
Kara bitkilerinin evrimsel yörüngesi boyunca, gametin azalmasına yönelik genel bir eğilim olmuştur. Tohumlu bitkiler bu eğilimi aşırıya götürür. Briyofitler gamet baskındır. Çekirdeksiz vasküler bitkiler sporofit baskındır, ancak gametofitleri çıplak gözle görülebilir. Tohumlu bitki gametofitleri ayrıca mikroskobik oranlara indirgenir. Tohumlu bitkilerdeki gametofitler sporangiadan gelişir ve sporofitik dokularda tutulur. Gametofitin sporofitik dokuya sarılması, gametofitin kurumasını önlediği için kara bitkileri için avantajlıdır. Tohumlu bitkiler yalnızca heterosporludur ve iki farklı spor türü üretir. Heterosporlu likofitlere benzer şekilde, tohumlu bitkiler oldukça azaltılmış gametofitlere sahiptir. Mikrosporangium, mikrosporositleri barındıran bir yapıdır. Diploid mikrosporositler, haploid mikrosporlar üreten mayoz bölünmeye uğrar. Mikrosporlar mitoza uğrar ve erkek gametofit veya mikrogametofite yol açar. Tohumlu bitkilerdeki mikrogametofit, polen olarak bilinen dört hücreli haploid bir yapıdır.
Likofitlerdeki megagametofitler, tümü bir megaspor duvarıyla kaplanmış besleyici doku ile çevrili sporofitik ( 2n ) bir embriyodan oluşur. Megasporangium, bir diploid megasporoktiye ( tohumlu bitkilerde megaspor ana hücresi olarak bilinir ) barındıran megasporagiumdan ( tohumlu bitkilerde nükleus olarak bilinir ) oluşan diploid, çok hücreli bir yapıdır. Nuclleus sonunda gelişmekte olan embriyo için besin rezervleri haline gelir. Tozlaşma, bir polen tanesi nüklele temas ettiğinde meydana gelir. Bu, megaspor ana hücresinin mayoz bölünmeye maruz kalmasına ve dört haploid megaspor üretmesine neden olur. Megasporlardan üçü yok olur ve geriye bir megaspor kalır. Hayatta kalan megaspor, dişi, haploid gametofit, megagametofit'e yol açmak için mitoz geçirir. Megagametofit, vazo şeklinde bir archegonium ile sarılmış bir yumurtayı barındıran çok hücreli bir yapıdır. Döllenme, sperm ve yumurta hücreleri kaynaştığında gerçekleşir.
Bazı likofitlerdeki megagametofitlerin aksine, megagametofitlerin ek bir dokusu, bir bütünlüğü vardır. Tohumlu bitkilerin bu modifiye megagametofitleri ovüllerdir. Bütünlük sert bir tohum kabuğuna dönüşür. Bu ekstra koruma, tohumların çimlenmeden önce uygun çevre koşulları ortaya çıkana kadar uykuda kalmasını sağlar. Megagametofitler geliştiren likofitlerin tohum kabuğu yoktur ve çimlenmeden önce uzun süreli uyku hali yapamazlar. Yumurta döllendikten sonra, nükleozdan gelen diploid besin rezervleri ile çevrili, megagametofitin içinde bulunan diploid sporofitik bir embriyoya dönüşür. Döllenmeden sonra, bütünlük nuclleus'u ve gelişmekte olan embriyoyu çevreler ve bir tohum kabuğu haline gelmek için sertleşir. Tohum filizlendikten sonra sporofitik ( 2n ) bir bitkiye veya sporofit haline gelir. Bitki cinsel olgunlaşmaya ulaştığında, yaşam döngüsünü yeniden başlatan mikro ve megasporangia geliştirir. Çamlarda bunlar kozalaklara yerleştirilir.
Kapalı tohumluların meyvelerin içine alınmış tohumları vardır
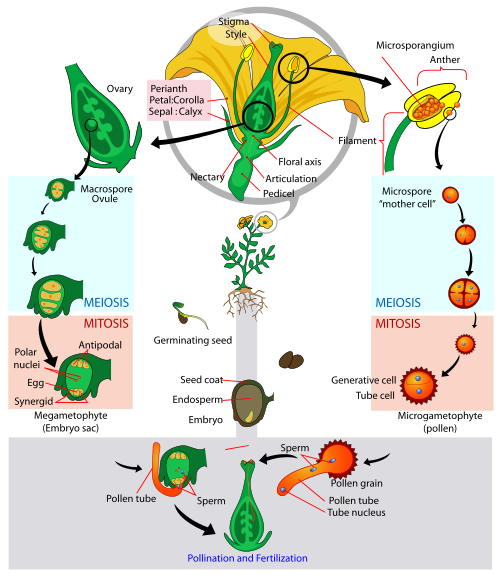
Kapalı tohumlular çiçekli bitkilerdir. Kapalı tohumlular şu anda kıtalardaki en baskın bitkilerdir ve monokotlar ( Sınıf Monocotyledonae ) ve dikotlar ( Sınıf Dicotyledonae ) ile temsil edilir. Tüm vasküler bitkiler gibi, kapalı tohumlular da sporofit baskındır. Gymnospermler gibi, kapalı tohumlular da heterosporlu, tohumlu bitkilerdir. Gymnosperm'den farklı olarak, kapalı tohumluların tohumları meyve olarak bilinen ek bir doku içerir. Meyvenin birincil işlevi tohum dağılımıdır. Meyveler hayvanlar tarafından yenebilir, hayvanların kürklerine veya tüylerine yapışabilir veya rüzgar veya su ile taşınabilir. Tohumların ana bitkiden uzağa dağılması, yavruların ana bitki ile tamamlanmamasını sağladığı için yavruların gelişmesi için faydalıdır. Gymnospermler gibi, kapalı tohumlular da haploid mikrosporlar üretmek için mayoz bölünmeye uğrayan mikrosporositler üretir. Bu mikrosporlar daha sonra mitoz yoluyla bölünür ve polen olarak bilinen iki hücreli mikrogametofitler üretir. Mikrosporositler, anter olarak bilinen bir yapı içinde yer alan bir mikrosporangium içinde tutulur. Çiçekler genellikle, çiçeğin tabanına tutturulmuş filamentlerin ( tipik olarak beyaz ) üzerinde tutulan tipik olarak sarı (bazen turuncu) olan polen üreten yapılar olan birçok anterlere sahiptir.
Gymnospermler gibi, kapalı tohumlular da ovül üretir. Bununla birlikte, gymnosperm ovüllerinin "çıplak" olduğu söylenir. Kapalı tohumlulardaki ovüller, yumurtalık olarak bilinen ek bir dokunun içine alınır. Meyveler yumurtalıklardan gelişir. Yumurtalıklar ( tipik olarak birçok ovül barındırır ), karpel olarak bilinen daha büyük bir yapının parçasıdır. Yumurtalığın üstünde, stil olarak bilinen bir boyun ( veya tüp ) uzanır. Stilin üstünde, stigma olarak bilinen ve poleni biriktirdikten sonra tutmak için yapışkan bir madde içeren bir organ bulunur. Yumurtalık içindeki ovüller, iki bütünleşme tabakası içinde yer alan bir megasporangium içerir. Megasporagium içindeki bir diploid megasporosit ( veya ana megaspor ), sadece biri hayatta kalan dört haploid megaspor üreten mayoz bölünmeye uğrar. Hayatta kalan megaspor mitoza uğrar ve embriyo kesesi olarak bilinen 8 hücreli bir dişi gametofit üretir. Embriyo kesesinin ekvatoruna en yakın iki hücre birleşerek bir diploid polar çekirdek oluşturur. Megagametofitin kalan hücreleri, embriyo kesesinin ekvatorundan uzaklaşır. Ovülün açıklığından en uzak üç hücre antipodaller olarak bilinirken, kalan üç hücre ( ovülün açıklığına en yakın ) bir yumurta hücresi ve iki sinerjik hücre ( veya "yardımcı" hücreler ) içerir.
Polen stigmaya ulaştığında, bir polen tüpü stilde büyür ve bütünlükler arasındaki yumurtalığa girer. Embriyo kesesinde iki sperm hücresi biriktirilir. Daha sonra, çift döllenme olarak bilinen kapalı tohumlulara benzersiz bir süreç gerçekleşir. Diğer tüm bitkiler gibi, bir sperm ve yumurta birleşerek bir embriyoya dönüşen diploid bir zigot oluşturur. İkinci sperm, diploid polar çekirdek ile birleşerek, endosperm olarak bilinen gelişmekte olan embriyo için bir besin rezerv dokusuna dönüşen bir triploid ( 3n ) hücre oluşturur. Döllenmeyi takiben, bütünlükler sertleşerek açık tohumlulara benzer şekilde sert bir dış tohum kabuğu oluşturur. Tohum filizlendikten sonra, bir kök kökü ortaya çıkar ve ardından kotiledonlar olarak bilinen bir ( monokotlar durumunda ) veya iki ( dikotlar durumunda ) tohum yaprağı gelir. Fide geliştikçe, endospermden gelen besin rezervleri ile desteklenir. Kapalı tohumlular aynı çiçekte ( hermafroditik ) organlarında ve pistillere, farklı çiçeklerde ( monokülerik ) organlarında ve pistillere veya ayrı erkek ve dişi bitkilerde ( ikievcik ) olabilir. Neredeyse tüm çiçekli bitkiler, ercik ve tabanca gelişiminin fiziksel veya zamansal olarak ayrılmasıyla kendi kendine tozlaşmayı azaltmak için mekanizmalar inşa etmiştir.